太赫兹在通信方向的应用 太赫兹通信和量子通信的区别
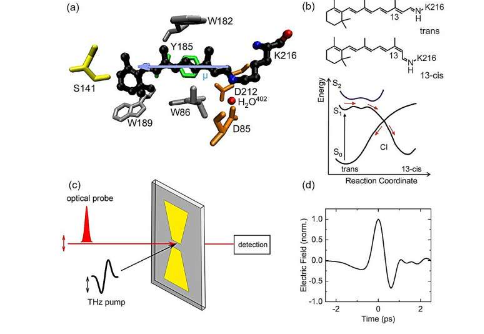
Rhodopsin细菌膜蛋白是一种质子泵,其中的质子传输是由光诱导的彩色视黄醇的异构化启动的。目前,参与这种超快速反应的分子量子态的特点是通过测量偶极矩。
太赫兹-斯塔克光谱学的新方法揭示了电子激发状态的混合物对光反应的路径和动力学的直接影响。
希芬视网膜质子是细菌类视网膜的头发颜色组,在吸收光线后其分子结构变化非常快。光激发将染色组提升到其激发电位能表面的特定范围,从中激发分子沿着反应坐标向激发状态和基本电位能表面的交点发展。激发状态的早期传播后,约500 fs=5&时间后激发;异构化发生在10 -13秒内通过此交叉点时。
到目前为止,控制反应动力学的激发电位能量的属性仍然存在争议。理论模型使用第一激发态 S 1 或混合量子态和第二激发态 S 2 的贡献。
这个问题需要一个新的实验理解的兴奋状态的特点。一个有前途的检测量是视网膜偶极矩,在基态S 0和第一和第二激发状态S 1和S 2明显不同。因此,光激发时偶极变化的测量应阐明与细菌视网膜的早期动力学相关的激发状态的特征。
(a) THz 斯塔克效应图。样品 E 位置的局部 THz 场导致能量从基本电子状态 S 0 和激发状态 S ex 偏移。因此,从S 0到S ex的光学过渡频率(垂直箭头)被改变. 符号和频率偏移的大小取决于S 0和S exµ状态的分子偶极矩;0 和 µex 投影到 E loc 方向。频率偏移和投影极性 Δ微型;关于µex - & micro;0 的乘积与本地字段 E loc 的乘积成正比。与视网膜偶极子在空间中随机定向的样品,实验采取平均为所有偶极子方向,从而导致 THz 诱导吸收光谱的增加。b) 在没有 THz 脉冲的情况下,最大 THz 电场值 (时间 t = 0) 和吸收光谱 A 0 (蓝线) 处的吸收变化 (符号) 和光频率之间的关系。THz脉冲引起瞬态光谱放大,A0的中心吸收减少,两侧吸收增加; 强黑线来自数字数据分析,光谱放大±12 THz, S 0 和 S ex & Delta 之间的偶极变化;微型;= 5 为 Debye。图片来源:MBI/T. Elsaesser
通过将一种新的太赫兹(THz)光谱学方法应用于斯塔克,来自柏林马克斯波恩研究所和洪堡大学以及慕尼黑马克西米利安路德维希大学的研究人员现在已经确定了视网膜电极在细菌光学红色中的变化(1 THz = 10 12 Hz = 10 12振荡每秒)。
正如他们在美国国家科学院院刊中报道的那样,光的激发导致视网膜偶极子的外观大约5 Debay(1.67×);10-29 库仑米) 的适度变化是远低于有序 S-1 兴奋状态的预期特征。
相反,他们的数据和理论分析表明 S2 状态与超高速激发状态动态前 120 fs 的平均时间混合解释了测量的偶极子的变化。这些结果支持在电子和核动力学在细菌视网膜早期存在明显混合的量子态的图表。
THz-Stark 光谱使用泵浦检测方法, 其中 1 皮秒(1 皮秒 = 10-12 秒)的 THz 泵浦脉冲提供强大的外部电场, 从视网膜的基态到激发状态的光学转变导致光谱 (Stark) 偏移。
这种偏移与基态和激发状态之间的偶极差相关 ∆&微;相称的。通过飞秒检测脉冲来测量样品的吸收变化,这些脉冲比THZ脉冲短,因此可以检测THZ场的瞬态效应。对于视网膜染色体的随机空间取向的样品, 可以观察到电子吸收带的光谱改善, 偶极子的变化 ≈ & 微;由此产生的。
随着时间的推移,放大跟随超短THZ脉冲的强度。在这个超短的时间尺度上,头发霜的蛋白质环境实际上是冻结和蛋白质动力学对实验观察值的影响可以忽略不计。
通过这种方式,THzStark光谱仪可以精确测量化学和生物学相关分子系统中的偶极矩。